




- -
- 100%
- +
Der englische Biomathematiker Ronald Fisher (1890 – 1962) kombinierte das Evolutionsmodell von Darwin und nachfolgende Beiträge von verschiedenen Genetikern zur einheitlichen Synthetischen Theorie der Evolution (oder Neodarwinismus). Er entwickelte damit die Grundlage|15◄ ►16| für quantitative Untersuchungen, welche auf Allelfrequenzen basieren (Fisher 1930). Evolution findet statt, wenn sich die Häufigkeiten von Allelen in einer Population verändern. Durch Mutationen können neue genetische Varianten entstehen. Durch Rekombination werden Allele in neuen Kombinationen an die Nachkommen weitergegeben.
Das zufällige Weitergeben von Allelen an die nächste Generation wird genetische Drift genannt. Dabei spielt die Populationsgröße eine wichtige Rolle. Das Allel-Set, das eine Generation an die Nachfolgegeneration weitergibt, ist statistisch gesehen immer eine Zufallsstichprobe. In großen Populationen werden auch Allele, welche in geringen Häufigkeiten vorkommen, an die nächste Generation weitergegeben, während bei kleinen Populationen seltene Allele verloren gehen. Durch genetische Drift kann sich die Nachfolgegeneration in den Allelhäufigkeiten deutlich von der Vorgängerpopulation unterscheiden.
Beim Gründereffekt besiedeln wenige Individuen einen neuen Lebensraum, etwa eine Insel. Diese Tiere repräsentieren nicht die ganze Bandbreite der in der Ausgangspopulation vorhandenen Allele. Wegen der geringen Vielfalt an mitgebrachten Allelen kann die neu entstandene Inselpopulation sich unter den veränderten Bedingungen anders entwickeln als die Ausgangspopulation. Dadurch erhöht sich die Wahrscheinlichkeit, dass auf der Insel eine neue Art entstehen kann (siehe: Wie entstehen neue Arten?).
Was ist eine Art?
Diese auf den ersten Blick triviale Frage ist gar nicht so leicht zu beantworten. In verschiedenen Artkonzepten wird mithilfe eindeutiger Kriterien versucht, die Vielfalt der Lebensformen auf unserem Planeten in diskrete Einheiten aufzuteilen. Da Evolution aber ein ständig fortschreitender Prozess ist, gibt es viele unterschiedlich stark ausgeprägte Übergangsformen zwischen Arten, die kaum durch eine gängige Definition taxonomisch zweifelsfrei abgegrenzt werden können. Die beiden am häufigsten verwendeten Definitionen für eine Art lauten:
• Eine Art ist eine Gruppe von Individuen, die sich in morphologischer, physiologischer oder biochemischer Hinsicht von anderen Gruppen unterscheidet (morphologische Definition einer Art, Morphospezies).
• Eine Art ist eine Gruppe tatsächlich und potenziell kreuzbarer Individuen, die sich mit Individuen anderer Gruppen unter natürlichen |16◄ ►17| Bedingungen nicht fortpflanzen (biologische Definition der Art, Biospezies).
Das Morphospezies-Konzept ist nützlich zur Unterscheidung der großen Zahl von lebenden, aber auch ausgestorbenen und nur fossil überlieferten Pflanzen- und Tierarten. Die folgenden Fakten zeigen aber die Grenzen dieses Konzeptes auf:
• Innerhalb einer Art können die Merkmale kontinuierlich variieren. Ein Phänotyp ist nicht vollständig durch den Genotyp determiniert, sondern ist das Ergebnis der Wechselwirkung von Genotyp und Umwelt. Ein und derselbe Genotyp kann je nach Lebens- und Umweltbedingungen unterschiedliche Formen und Größen bewirken. Viele Arten zeichnen sich durch eine hohe phänotypische Plastizität aus. So variiert beispielsweise die Blattform und Größe des Löwenzahns (Taraxacum officinale) stark in Abhängigkeit von der Niederschlagsmenge, Sonneneinstrahlung und Jahreszeit zum Zeitpunkt der Blattbildung.
• Innerhalb einer Art können Merkmale diskret variieren (intraspezifischer Polymorphismus). Individuen der Hain-Bänderschnecke (Cepaea nemoralis) weisen eine gelbe, rosa oder braune Gehäusefarbe auf. Zusätzlich können die Gehäuse mit einem bis fünf, manchmal zusammenhängenden dunkelbraunen Bändern verziert sein. Alle diese Merkmale sind genetisch determiniert (Murray 1975). Das sehr variable Aussehen der Hainbänderschnecke verleitet zu einer Zuordnung von Individuen in verschiedene Arten.
• Viele Arten durchlaufen während der Individualentwicklung verschiedene Stadien (z.B. Larvenstadien bei Fliegen, Raupen und Puppe bei Schmetterlingen). Larvenstadien lassen sich oft nur mit großen Schwierigkeiten einer bestimmten Art zuordnen. Es ist auch vorgekommen, dass Larven einer schon bekannten Art als eigene, neue Art beschrieben wurden.
• Bei zahlreichen Arten sehen weibliche und männliche Individuen unterschiedlich aus (z.B. Stockente, Auerhuhn, Rothirsch, zahlreiche Insekten). Dieser sogenannte Sexualdimorphismus hat in verschiedenen Fällen dazu geführt, dass Weibchen und Männchen derselben Art ursprünglich verschiedenen Arten zugeordnet wurden.
• Biologisch völlig verschiedene Arten können aufgrund ähnlicher Selektionsbedingungen in ihrem Phänotyp konvergieren, sodass sie rein äußerlich kaum mehr zu unterscheiden sind.
|17◄ ►18|
Das biologische Artkonzept basiert auf der Annahme, dass die Isolationsmechanismen zwischen den einzelnen Arten auf biologischen Eigenschaften der Organismen beruhen. Zwischenartliche Kreuzungen werden durch ethologische, morphologische, physiologische oder genetische Isolationsmechanismen verhindert. Dadurch bilden die Individuen einer Art eine Fortpflanzungsgemeinschaft mit einem gemeinsamen Genpool. Aber auch das biologische Artkonzept hat seine Grenzen:
• Arten, die sich nur ungeschlechtlich vermehren (klonal oder durch Parthenogenese), werden durch die Definition des biologischen Artkonzeptes nicht erfasst. Zu diesen gehören alle Prokaryoten (Mikroorganismen), einige Pilze und Pflanzen, verschiedene Insekten und Echsen.
• Viele Pflanzen- und Tierarten kreuzen sich auch in der Natur (z.B. Steinkorallen, Orchideen). Nach dem biologischen Artkonzept sind einige Orchideenarten keine getrennten Arten.
• Individuen, die sich nie in einer Population fortpflanzen können (z.B. Arbeiterinnen bei Bienen oder Ameisen) werden vom biologischen Artkonzept nicht erfasst, obwohl sie zum Genpool beitragen.
Trotz dieser Einschränkungen findet das biologische Artkonzept als «vorstellbares Modell» häufig Verwendung in der Ökologie und Naturschutzbiologie.
In neuerer Zeit werden immer mehr molekularbiologische Methoden zur Unterscheidung einzelner Arten eingesetzt (Exkurs: DNA-Taxonomie und DNA-Barcoding). DNA-Taxonomie ermöglicht die Entdeckung und Beschreibung neuer Arten, ist aber kein Ersatz für die klassische Taxonomie. DNA-Barcoding funktioniert nicht ohne die Taxonomie- und Systematik-Fachleute der einzelnen Organismengruppen. Entscheidende Fortschritte in der Biodiversitätserfassung wird es nur durch den komplementären Einsatz von klassischen und molekularen Methoden geben.
|18◄ ►19|
DNA-Taxonomie und DNA-Barcoding
Zur Unterscheidung einzelner Arten werden zunehmend molekularbiologische Methoden (vor allem Sequenzierung geeigneter DNA-Abschnitte) angewendet (Tautz et al. 2003). Bisher wurden DNA-Sequenzen meistens als Zusatzkriterien zur sicheren Artbestimmung angewendet. Seit geraumer Zeit gibt es aber Bemühungen, alle bekannten Arten auch molekularbiologisch zu charakterisieren. Dabei wird nicht das ganze Genom sequenziert, sondern nur Abschnitte (oft die Cytochrom-Oxidase Untereinheit I).
DNA-Barcoding ermöglicht die Identifizierung von bereits bekannten Arten mittels eines arttypischen DNA-Fragmentes durch den Abgleich mit einer bestehenden Datenbank (www.barcodinglife.com). Das funktioniert ähnlich wie das Barcoding (Streifencode) auf Verkaufswaren im Supermarkt, nur dass hier nicht schwarze Streifen, sondern DNA-Basenabfolgen zur Identifizierung dienen (Streit 2007). Sequenziergeräte dürften in Zukunft kleiner und preiswerter werden, was charakteristische Zuordnungen von DNA-Proben zu konkreten Taxa vermutlich sogar im Freiland ermöglichen wird. Bis zur Etablierung einer weltweit auch nur annähernd vollständigen Barcoding-Datenbank ist der Arbeitsaufwand allerding sehr groß.
Wie entstehen neue Arten?
Der entscheidende Schritt einer potenziell neuen Art erfolgt mit der vollständigen Trennung des Genpools einer Population von anderen Populationen. Dies ist der Fall, wenn sich die Individuen der betrachteten Population nicht mehr mit den Individuen der anderen Populationen kreuzen. Ist der Genpool einer Teilpopulation einmal isoliert, folgt er einer eigenen, unabhängigen Entwicklung, die durch natürliche Selektion, genetische Drift und Mutation beeinflusst wird. Verschiedene Prozesse können zur Artbildung führen:
• Allopatrische oder geografische Artbildung,
• sympatrische Artbildung (Polyploidie und Artbildung durch Konkurrenz).
Allopatrische (geografische) Artbildung kann eintreten, wenn das Verbreitungsgebiet einer Art durch äußere Prozesse in zwei oder mehr Teile aufgespalten wird. Eine Population wird durch die Neubesiedlung einer
|19◄ ►20|
Insel, durch Austrocknung von Gewässern, durch Vergletscherung, Vulkanausbruch oder durch Landhebung vom Hauptverbreitungsgebiet der Art abgetrennt. Zunächst wird sich der Genpool von der isolierten Population kaum vom Genpool der Art im Hauptverbreitungsgebiet unterscheiden. Falls es aber Unterschiede in den klimatischen Bedingungen und anderen selektiven Faktoren zwischen dem abgetrennten Gebiet und dem Hauptgebiet gibt, werden sich die Genfrequenzen durch natürliche Selektion, genetische Drift und Mutation verändern. Das abgetrennte Gebiet ist beispielsweise wärmer und beherbergt eine andere Prädatorart. Dann wird die natürliche Selektion diejenigen Individuen bevorzugen, die am besten an ein wärmeres Klima angepasst sind und den neuen Prädatoren ausweichen können. Aufgrund der unterschiedlichen Selektionsdrucke auf die beiden Populationen werden sie sich nun langsam genetisch auseinanderentwickeln. Es können Veränderungen im Verhalten, in der Physiologie und der Morphologie zwischen den beiden Populationen entstehen, die keine erfolgreiche Fortpflanzung mehr zulassen und somit zur reproduktiven Isolation führen.
Es gibt zahlreiche Beispiele von allopatrischer Artbildung. In den bewaldeten Bergketten der Insel Oahu (Hawaii) konnten 41 endemische Baumschneckenarten der Gattung Achatinella nachgewiesen werden (Hadfield 1986). Getrennt durch Bergrücken beherbergte jedes Tal ihre eigene Art von diesen Baumschnecken, die sich von auf Blättern wachsenden Pilzen ernährten. Durch Zerstörung des Lebensraumes, dem Aussetzen von nicht-einheimischen Schneckenprädatoren und übermäßige Sammeltätigkeiten sind in den letzten Jahrzehnten aber über 30 Arten ausgestorben (Kapitel 8).
In den Alpen isolierte die wiederholte Vergletscherung während der Eiszeiten zahlreiche Pflanzen- und Tierarten auf wenigen eisfreien «Inseln». In Perioden zwischen den Eiszeiten konnten sich manche Restpopulationen nur beschränkt wieder ausbreiten. Viele Populationen entwickelten sich während der Isolation zu neuen Arten. Allein in den österreichischen Alpen kommen 103 endemische Pflanzenarten und -unterarten vor, die auf diese Weise entstanden sein dürften (Essl et al. 2009; siehe Exkurs «Endemische Arten», Seite 32).
Sympatrische Artbildung erfolgt ohne geografische Trennung. Im Pflanzenreich ist sympatrische Artbildung durch Chromosomenverdoppelung oder -vervielfachung (Polyploidie) häufig. Durch Unregelmäßigkeiten in der Chromosomenpaarung oder -verteilung während der Reduktionsteilung (Meiose) können die Nachkommen doppelte Chromosomensätze haben. Eine Kreuzung von Individuen mit einer |20◄ ►21| unterschiedlichen Anzahl von Chromosomensätzen führt in der Regel zu sterilen Hybriden, die keine normale Meiose mehr durchführen können. Ein polyploider Organismus hat somit über die Verdoppelung des Chromosomensatzes einen reproduktiven Isolationsmechanismus gegenüber den ursprünglichen Individuen aufgebaut. Im Vergleich zur allopatrischen Artbildung ist Polyploidie ein sehr schneller Artbildungsprozess.
Bei Autopolyploidie stammen die Gameten von der gleichen Elternart. Autopolyploidie ist beispielsweise beim Mittleren Wegerich (Plantago media; Populationen mit Chromosomenzahl 12 und 24) gut untersucht. Bei Allopolyploidie stammen die Gameten von verschiedenen Elternarten, die häufig nahe verwandt sind. Allopolyploidie spielt bei der Entstehung von vielen Kulturpflanzen eine wichtige Rolle. Bei Tieren ist Polyploidie seltener, kommt aber bei Arten vor, die sich durch Parthenogenese fortpflanzen, beispielsweise beim Salinenkrebs (Artemia salina).
Sympatrische Artbildung kann auch als Ergebnis einer disruptiven Selektion auftreten, die zwei oder mehr Phänotypen innerhalb einer Population begünstigt. Wenn unterschiedliche Eigenschaften ähnlich gut geeignet sind, um Nahrungsquellen zu erschließen (beispielsweise zwei verschiedene Schnabelformen) oder andere Bedürfnisse zu erfüllen, kann natürliche Selektion auch unterschiedliche Phänotypen innerhalb einer Population begünstigen. Eine Voraussetzung ist allerdings, dass sich Organismen nur mit dem gleichen Phänotyp paaren (assortative mating). Diese Form der Artbildung konnte bisher allerdings nur in wenigen Studien nachgewiesen werden, eine davon bezieht sich auf Landschnecken mit rechts- und linksgewundenen Gehäusen.
Die Entstehung neuer Arten ist normalerweise ein langsamer Prozess, der Hunderte bis Tausende von Generationen erfordert (Ausnahme: Polyploidie). Die Evolution neuer Gattungen und Familien verläuft sogar noch langsamer. Generell ist die Artbildungsrate auf Inseln größer als auf dem Festland. Dies ist unter anderem auf die Isolation der Inseln, den Gründereffekt (siehe oben) und das Fehlen gewisser Selektionsfaktoren zurückzuführen (fehlende Prädatoren, reduzierte Konkurrenz). Ein klassisches Beipiel für Artbildung auf Inseln sind die Darwin-Finken auf den Galapagos-Inseln.
Eine neuentdeckte Form von Lebewesen gilt als wissenschaftliche Art, wenn sie nach den geltenden Nomenklaturregeln beschrieben wurde.
|21◄ ►22|
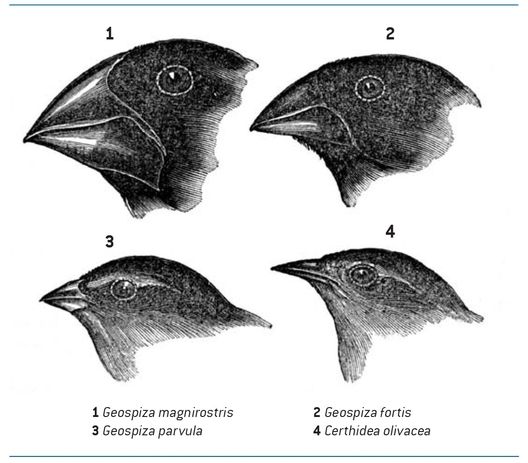
Abb. 1: Adaptive Radiation bei den Darwin-Finken auf den Galapagos-Inseln. Die unterschiedlichen Schnabelformen sind Anpassungen an verschiedene Nahrungsweisen. Alle Arten gehen auf eine einzige Stammart zurück (aus Darwin 1845, mit Bewilligung von van Wyhe J. (ed.) The Complete Work of Charles Darwin Online, www.darwin-online.org.uk).
|22◄ ►23|
Binäre (binomiale) Nomenklatur und Artbeschreibung
Carl von Linné (lateinisch Linnaeus; 1707 – 1778), ein schwedischer Arzt und Naturwissenschaftler, entwickelte das System der binären Nomenklatur der Pflanzen und Tiere. Der zweiteilige Name setzt sich zusammen aus dem Namen der Gattung, der stets als Nomen mit einem Großbuchstaben beginnt, und einem kleingeschriebenen Epitheton (häufig ein Adjektiv), welches in Kombination mit der Gattung die Art charakterisiert. Jede solche Kombination von zwei Namen darf nur einmal – also nur für eine Art – vergeben werden. Diese Namen entstammen gewöhnlich der lateinischen oder griechischen Sprache. Nichtlateinische Namen werden lateinisiert. So setzt sich beispielsweise der wissenschaftliche Name der Amsel aus den lateinischen Bezeichnungen turdus (Gattung Echte Drosseln) und merula zusammen und lautet vollständig Turdus merula Linnaeus, 1758. Die zur gleichen Gattung gehörende Singdrossel heisst Turdus philomelos Brehm, 1831. In der wissenschaftlichen Literatur wird der zweiteilige Artname in kursiver Schrift dargestellt, oft gefolgt vom Autorzitat, d.h. dem Namen (oder Namenskürzel) der Person, welche die erste gültige wissenschaftliche Beschreibung der Art verfasst hat. Darauf folgt noch das Jahr der Veröffentlichung dieser Beschreibung. In der Systematik werden die Gattungen in die nächsthöhere Stufe der Familie zusammengefasst und verschiedene Familien wiederum in die nächsthöhere Stufe der Ordnung. Eine Klasse umfasst verschiedene Ordnungen.
Zur Beschreibung und Benennung von Arten wurden spezielle, weltweit geltende Nomenklaturcodes entwickelt, die sich zwischen Pflanzen (www.ibot.sav.sk/icbn) und Tieren (www.iczn.org) geringfügig unterscheiden. Durch diese Regeln sollten wiederholte Beschreibungen der gleichen Art sowie der Gebrauch desselben Namens für mehr als eine Art vermieden werden. Ein wichtiger Bestandteil einer Artbeschreibung ist das Festlegen eines Referenzexemplars oder -musters (Holotyp oder Holotypus). Das Typus-Exemplar muss in einem Museum aufbewahrt werden, damit es zu Vergleichszwecken zugänglich ist.
|23◄ ►24|
Adaptive Radiation bedeutet die Auffächerung einer wenig spezialisierten Art in viele stärker spezialisierte Arten. Dies erlaubt beispielsweise die Nutzung eines breiteren Spektrums von Nahrungsressourcen. Bekannte Beispiele für die Radiation von Tiergruppen sind wiederum die Darwin-Finken mit ihren unterschiedlichen Schnabelformen (Abb. 1, S. 22) sowie die Kleidervögel auf Hawaii, die Buntbarsche in den großen Seen von Afrika und die marinen Kegelschnecken (Conidae). Diese Schnecken sind hochspezialisierte, nachtaktive Prädatoren, die zum Nahrungserwerb eine Art Harpune benutzen, welche sich aus einem Zahn der Radula (Raspelzunge) entwickelt hat. Durch den nadelspitzen, hohlen Zahn injiziert sie ein Gift ins Beutetier. Es gibt rund 600 rezente Arten von Kegelschnecken. Viele fressen Borstenwürmer; andere sind aber auf Mollusken spezialisiert, wiederum andere jagen ausschließlich Fische oder erbeuten Krabben, denen sie im Sand vergraben auflauern. Alle töten ihre Beute aber mit Gift.
Weiterführende Literatur
Darwin C. (1859) (Übersetzung 1981) Über die Entstehung der Arten. Reclam, Stuttgart.
Steinke D. & Brede N. (2006) DNA-Barcoding: Taxonomie des 21. Jahrhunderts. Biologie unserer Zeit 36: 40 – 46.
|24◄ ►25|
2
Biodiversität verändert sich
Zusammenfassung
Im Laufe der Erdgeschichte entwickelten sich aus einzelligen mehrzellige Organismen und später komplexe Lebewesen. Zahlreiche ursprünglich aus dem Meer stammende Organismengruppen besiedelten das Festland. Generell nahm die gesamte Artenvielfalt auf der Erde mit der Zeit zu, wurde aber durch mehrere Massenaussterbeereignisse zwischenzeitlich wieder reduziert. Momentan erleben wir das größte Massenaussterben, welches je in der Erdgeschichte stattgefunden hat. Im Gegensatz zu den früheren Aussterbeereignissen ist dieses Mal eine einzige Art der Verursacher, nämlich der Mensch. In diesem Kapitel wird auch gezeigt, wie schwierig eine präzise Ermittlung der Zahl der tatsächlich auf der Erde existierenden Arten ist. Zurzeit sind rund 1,8 Millionen Arten bekannt. Viele Gruppen sind bisher aber äußerst lückenhaft erfasst worden. Die Gesamtzahl der Mikroorganismen, Pilze, Pflanzen und Tiere auf der Erde wird auf 10 – 20 Millionen Arten geschätzt.
Artenvielfalt im Laufe der Erdgeschichte
Der zeitliche Ablauf der Evolution kann anhand von Fossilien rekonstruiert werden. Entsprechend der vertikalen Aufeinanderfolge fossilführender Gesteinsschichten lassen sich Gesellschaften vorzeitlicher Lebewesen in eine zeitliche Reihenfolge bringen und mithilfe radiometrischer Methoden kann das Alter der Gesteine bestimmt werden. Das Vorkommen und Alter gewisser Fossilien geben zudem Hinweise, wann im Verlauf der Stammesgeschichte bestimmte Innovationen und Aufsplitterungsereignisse auftraten. Bestimmte gemeinsame Merkmale in Gruppen von Lebewesen legen nahe, dass sie ursprünglich von einer Art (oder einem Genpool) abstammen. Mit verschiedenen Ansätzen (vergleichende Anatomie, Morphologie, molekulare Techniken) können Verwandtschaftsverhältnisse sowohl zwischen Arten als auch zwischen taxonomischen Gruppen untersucht werden. Mithilfe molekularer |25◄ ►26| Schätzungen (molecular clock) lassen sich die Zeitpunkte gewisser Ereignisse auch berechnen.
Das Alter der Erde wird auf 4,5 Milliarden Jahre geschätzt. Die frühesten Fossilien von lebenden Organismen – einfache Bakterien – stammen aus ca. 3,5 Milliarden altem Gestein. In den nächsten 3 Milliarden Jahren entfaltete sich das Leben vorwiegend im Wasser. Die ersten Eukaryoten entstanden vermutlich vor 2 Milliarden Jahren. Aus Einzellern entwickelten sich Zellkolonien und schließlich mehrzellige Organismen.
Die Landbesiedelung, d.h. die Anpassungen von aquatischen Lebewesen an eine terrestrische Lebensweise, fand wiederholt und unabhängig voneinander in verschiedenen Gruppen wie Einzellern, Pilzen, Pflanzen, Schnecken und Wirbeltieren statt (Little 1990). Durch fossile Belege relativ gut dokumentiert ist die Landbesiedelung der Pflanzen vor ca. 460 – 480 Millionen Jahren (im Devon; Tabelle 2). Die Urfarne traten vor etwa 400 Millionen Jahren auf, erreichten schnell eine große Vielfalt, starben aber später wieder aus. Andere Farnarten übernahmen ihre Nischen. Die Gymnospermen (Nacktsamer) wurden erstmals in Gesteinen des Devons gefunden und wiesen während des Mesozoikums eine größere Vielfalt auf als heute. Die Gymnospermen erlebten in den letzten 100 Millionen Jahren einen Rückgang, als die Angiospermen (Bedecktsamer) eine große Artenvielfalt entwickelten. Heute gibt es etwa 800 Gymnospermen-Arten, aber rund 240 000 Angiospermen-Arten. Durch fossile Belege gut abgesichert ist die Landbesiedelung der Wirbeltiere (Tetrapoden) während des Devons, d.h. der evolutionäre Übergang von Fischen zu den Vorformen der heutigen Amphibien (Tabelle 2, S. 28 – 29).
Bei der Betrachtung der Evolution verschiedener Organismengruppen muss auch berücksichtigt werden, dass die Landmasse (die späteren Kontinente) im Verlauf der Erdgeschichte sich in der Lage auf der Erdkugel, in der Größe (durch Schwankungen im Meeresspiegel) sowie im Ausmaß der Isolation einzelner Teile verändert hat (Abb. 2). Im Zeitraum, als die Landbesiedelung von zahlreichen Gruppen stattfand, gab es eine einzige Landmasse (Pangaea). Später, als die Dinosaurier verbreitet waren, trennte sich Pangaea durch plattentektonische Kräfte in Laurasien und Gondwana auf. Die nachfolgende Isolation der Kontinente trug wesentlich zur Entwicklung einzelner Gruppen bei. Ein gutes Beispiel dafür ist der Inselkontinent Australien. Nach der Auflösung Gondwanas in die einzelnen Erdteile driftete die australische Platte lange Zeit weitgehend isoliert in östlicher Richtung. Ein Kontinente übergreifender Artenaustausch konnte nicht mehr stattfinden. In der Isolation Australiens|26◄ ►27| konnten so Organismengruppen überdauern, die sonst fast überall ausgestorben sind, wie etwa die Kloakentiere (Protheria). Auch die Beuteltiere (Metatheria) konnten hier überleben und entwickelten sich zu einer artenreichen Gruppe, während es mit Ausnahme weniger Arten in Amerika auf der Erde sonst keine Vertreter dieser einstmals weit verbreiteten Gruppe mehr gibt.
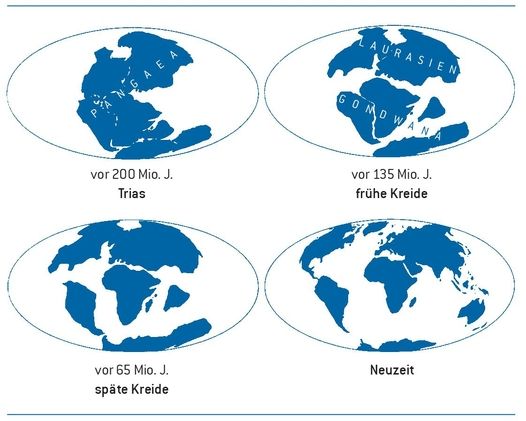
Abb. 2: Anordnung der Landmasse in der Trias, frühen und späten Kreidezeit sowie in der Neuzeit (nach Cloud 1978).
Fossilien belegen die Zu- und Abnahmen in der Diversität einzelner Gruppen. Neue Arten haben sich entwickelt, viele sind auch wieder ausgestorben. In Organismengruppen, über die aufgrund von Fossilfunden ausreichende Erkenntnisse vorliegen, ist die Artenzahl seit ihrem ersten Auftreten meistens angestiegen. Die heute lebenden Pflanzen- und Tierarten machen – je nach Schätzung – weniger als 1 % bis maximal 4 % der Arten aus, die jemals auf der Erde gelebt haben. Das Aussterben einer Art ist demnach ein fast ebenso häufiges Ereignis in der Erdgeschichte wie das Erscheinen einer neuen. Es gab aber auch Phasen der Abnahme der Diversität. Aufgrund der Fossilienfunde konnten in der Vergangenheit mindestens sechs Aussterbeereignisse, die sich innerhalb bestimmter Erdepochen auf relativ kurze Zeitabschnitte konzentrierten, dokumentiert werden (Abb. 3). Ein solches Massenaussterben ereignete sich beispielsweise am Ende des Perms vor rund 250 Millionen Jahren. Damals verschwanden in den marinen Flachwasserbereichen rund 90 % aller Wirbellosen. Ein weiteres Massenaussterben gab es am Ende der Kreidezeit vor rund 65 Millionen Jahren, als die Dinosaurier verschwanden. Heute befinden wir uns mitten in einem weiteren Massenaussterben. Es wird geschätzt, dass im 21. Jahrhundert zwischen 10 000 und 25 000 Arten jährlich auf der Erde aussterben; dies entspricht ein bis drei Arten pro Stunde. Das Artensterben verläuft gegenwärtig mindestens tausend Mal schneller als jemals zuvor in der Erdgeschichte. Zudem wird im Unterschied zu den früheren Ereignissen das jetzige Massenaussterben durch eine einzelne Art verursacht, nämlich durch den Menschen. Die Ursachen für das zurzeit stattfindende Massenaussterben werden detailliert in Kapitel 8 behandelt.